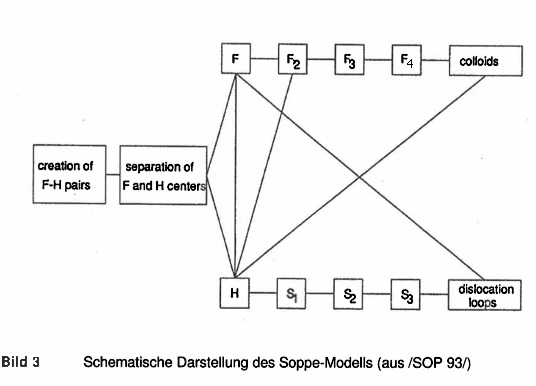
Soppe-Modell -oben- und DTVH-Modell -unten-. Im Vergleich zum DTVH-Modell fehlt beim Soppe-Modell der mittlere Reaktionspfad und die daran anschließende Poren- und Rißbildung (rechts in Kreis und Ellipse).
DTVH-Modell (Quelle: Fig. 2.2 in Sugonyako, 2007) |
Bild 3: Mechanismen im Soppe-Modell (SOP 93 und Soppe et al. 1996, siehe auch linke Spalte "Jain-Lidiard" in Tabelle 2.1). Beta- und Gammastrahlung regen das NaCl-Gitter zu Excitonen an. Beim strahlungsfreien Zerfall eines Excitons wird als Folgeschaden ein F- und H-Zentrum-Paar gebildet.
F-Zentren können sich bei ihrer Brownschen Bewegung ("thermischen random walk") zu 2er-, 3er- und schließlich 4er-Gruppen zusammenballen (F2, F3, F4, "self-trapping"). F4-Zentren gelten als "homogene" Keime für Na-Kolloide (es gibt darüberhinaus auch heterogene Kolloidkeime im NaCl-Gitter). H-Zentren können sich -ähnlich wie F-Zentren- zusammenballen ("cluster") oder werden an Gitterverunreinigungen eingefangen und verbinden sich mit diesen zu Komplexen: S1-, S2-, S3-Komplexe sind Verbindungen mit 1, 2 oder 3 H-Zentren. Das S3-Zentrum gilt als Keim für einen Stufenring ("dislocation loop", Bildung eines dislocation loop's im Zeichentrickfilm): Reihe 1 im Jain-Lidiard-Modell in Tabelle 2.1 Zwei H-Zentren in der Nachbarschaft einer Stufenversetzung bilden ein molekulares Cl2-Zentrum, indem sie ein NaCl-Molekül aus dem regulären NaCl-Gitter "ausgraben". Dieses NaCl-Molekül wandert zur zusätzlichen Ebene der Stufenversetzung, was zu einem vollständigen Stufenring führt: Reihe 2 im Jain-Lidiard-Modell in Tabelle 2.1. Stufenversetzungslinien ("edge dislocations") wirken als nicht sättigbare Senken für F- und H-Zentren, aber wegen der stärkeren elastischen Wechselwirkung an diesen Linien binden sich die H-Zentren dort fester als die F-Zentren. Diese zu Ungunsten der F-Zentren ausgehende Konkurrenz zwischen F- und H-Zentren um Bindung an die Stufenversetzungslinien führt einerseits zur Bildung von Cl2-Blasen an den Stufenversetzungen und andererseits zu einem Überschuß an F-Zentren irgendwoanders im Gitter und letztendlich -wie oben beschrieben- zu Na-Kolloiden: Reihen 3 bis 5 im Jain-Lidiard-Modell in Tabelle 2.1. Begrenzung des Kolloidwachstums: Reihe 7 im Jain-Lidiard-Modell in Tabelle 2.1. Das Kolloidwachstum
Graphisch dargestellt: Im Soppe-Modell fehlt der entsprechende Reaktionspfad des DTVH-Modells, dargestellt als mittlerer Pfad in Fig. 2.2 und in der rechten Spalte der Tabelle 2.1, in dem Poren bei ihrem Wachstum kolloidales Natrium und Cl2-Blasen in sich aufnehmen.
In Ergänzung dazu schlagen Dubinko und Mitarbeiter
"einen Mechanismus für das Wachstum von Poren infolge einer Ballung von Doppel-Leerstellen vor [Doppel-Leerstelle (Leerstellenpaar) = 1 Leerstelle im Na+-Untergitter plus 1 benachbarte Leerstelle im Cl--Untergitter], welche durch Wechselwirkung zwischen Stufenversetzungen und Excitonen erzeugt werden. Der Prozess ist folgender:
|
Das ... Schicksal der über den Radiolyseprozeß entstandenen primären Defekte, d. h. der F- und H-Zentren, ist von einer Reihe von Faktoren abhängig. Aufgrund der Translationssymmetrie des Kristallgitters kann ein Defekt eine große Anzahl gleichwertiger Gitterpositionen einnehmen, die durch Energiebarrieren getrennt sind, die durch thermische Gitterschwingungen überwunden werden können. Die Beweglichkeiten der beiden primären Defekte sind sehr unterschiedlich.
Die Wanderungsenthalpien für Leerstellen im Kationenuntergitter, Vc, bzw. für Leerstellen im Halogeniduntergitter, Va, betragen beim NaCI 0,69 bzw. 0,77 eV, und liegen für F- Zentren in derselben Größenordnung. Sie können daher erst bei Raumtemperatur oder höheren Temperaturen wandern.
Zwischengitteratome bzw. -moleküle, in deren Umfeld die elastische Gitterverzerrung wesentlich größer ist, sind dagegen erheblich beweglicher und beginnen bereits bei Temperaturen um 30 K zu wandern.
Bei seiner Wanderung durch das Kristaiigitter kann ein Defekt auf einen anderen räumlich getrennten primären Defekt oder auf lokalisierte strahleninduzierte bzw. ursprünglich im Kristall vorhandene Defekte treffen und mit diesen in Wechselwirkung treten. Dies führt zur Verringerung von freien primären Defekten. Diese Prozesse laufen selbst bei moderaten Temperaturen diffusionskontrollert ab.
Das weitere Schicksal der primären Defekte hängt davon ab, welche Reaktionspartner bei der Diffusion durch den Kristall angetroffen werden.
Trifft ein H-Zentrum auf ein F-Zentrum, rekombinieren beide Defekte unter gegenseitiger Auslöschung mit Energiefreisetzung miteinander, wobei wieder das ursprüngliche Kristallgitter ausgebildet wird ["Defektausheilung"].
Treffen dagegen zwei gleichartige Defekte zusammen, führen Austauschwechselwirkungen zu einer Bindung zwischen den Defekten. So besitzen zwei F-Zentren, die zwei benachbarte Plätze im Anionengitter besetzen, eine Bindungsenergie von ca. 0,5 - 1,0 eV. Durch Anlagerung weiterer Defekte an solche Defektpaare bilden sich größere Defektaggregate. Im Falle der F-Zentren führt dies zur Bildung von Natrium-Kolloiden,
die H-Zentren lagern sich zu zunehmend komplexen Polyhalogenen zusammen. Zwei H-Zentren bilden ein neutrales Cl2-Molekül oder ein Cl3--Quasimolekül, vier H-Zentren bilden X5- usw. bis der Defektcluster so groß wird, daß er als Gasblase aus (Cl2)n betrachtet werden kann. Dieser Vorgang läuft an Stufenversetzungen (Stufenringen) ab, die dadurch vergrößert werden.
2.3 Mathematische Modelle zur Beschreibung der Strahlenschädenbildung
Zur mathematischen Beschreibung des Einflußes der Parameter gamma-Dosisleistung, Temperatur, Verunreinigungen usw. auf die Bildung von Defektaggregaten sind verschiedene Ansätze entwickelt worden. Die Theorien können in drei allgemeine Kategorien eingeordnet werden. [Im Folgenden wird nur die eine, der makroskopische Ansatz beschrieben.]
Ein makroskopischer Ansatz basiert auf kinetischen Gleichungen, die die Änderungen der makroskopischen Konzentrationen der primären Defekte und ihrer Aggregate als Funktion der Zeit beschreiben. Die Konzentrationen werden über das gesamte Volumen des Kristalls gemittelt und hängen nicht von der Position der Defekte ab. Auf diesem Ansatz basieren die Jain-Lidiard-Theorie (JL-Theorie) /JAI 77, LID 79/ und die von den Hartog und seinen Mitarbeitern /SEI 92/ sowie von Soppe /SOP 93, SOP 94b/ vorgenommenen Erweiterungen. Die prinzipielle Schwäche der makroskopischen Ansätze liegt in der Verwendung phänomenologischer Parameter, besonders Geschwindigkeitskonstanten, die als zeit- und konzentrationsunabhängig angesetzt werden. Ferner werden mehrere Parameter benötigt, deren Werte bisher weder experimentell noch theoretisch genau bestimmt worden sind [siehe auch Gl. 2.2 - 2.5 in Bergsma & Heijboer, 1983]. Trotz dieser Einschränkungen sind diese Ansätze recht erfolgreich in der Beschreibung vieler wesentlcher Aspekte der Radiolyseprozesse in Alkalihalogeniden und insbesondere im NaCI.
...
Bisher sind die auf dem makroskopischen Ansatz basierenden Theorien hinsichtlich der Richtigkeit der Modellvorhersagen am erfolgreichsten. Diese Modelle sind im Statusbericht von 1993 /GIE 93/ ausführlich beschrieben worden. Hier wird kurz auf das Soppe-Modell eingegangen, da damit die Modellrechnungen mit realistischen Randbedingungen durchgeführt wurden, die im Kap. 4 [von GRS-142] beschrieben werden.
Das Soppe-Modell stellt eine Weiterentwicklung der ursprünglichen JL-Theorie dar, da die JL- Theorie nicht richtig den Einfluß von größeren Konzentrationen an Verunreinigungen und Stufenversetzungen auf das Kolloidwachstum beschreiben kann. Außerdem wird die Nukleationsphase für das Kolloidwachstum nicht modelliert, sondern über einen zu einfachen Ansatz berücksichtigt. Das Soppe-Modeii ähnelt dem Modell von Groote und Weerkamp ISEI 92/, unterscheidet sich von diesem aber in einigen wesentlichen Details. Es weist folgende Merkmale auf [s. Bild 3]:
Einfang der F-Zentren und Kolloidwachstum. F-Zentren lagern sich zusammen, wobei sukzessive F2-, F3- und F4-Zentren gebildet werden. Die F4-Zentren werden als Keim für das Kolloidwachstum durch homogene Nukleation betrachtet. Zusätzlich wird eine bestimmte Anzahl von heterogen erzeugten Keimen durch Anlagerung von F-Zentren an andere Stellen angenommen. Die Aggregation von F- Zentren zu Kolloiden erfolgt über ungerichtete Diffusion (random-walk).
Einfang von H-Zentren durch Verunreinigungen. Dieser Vorgang führt zur Bildung von S1, S2 und S3-Komplexen, die durch sukzessive Anlagerung von H-Zentren entstehen. Das S3-Zentrum wird als Beginn einer neuen Stufenversetzungslinie betrachtet.
Wachstum der Dislokationsstufen und Bildung von molekularem Chlorgas. Dislokationen reagieren mit F- und H-Zentren, wobei allerdings die Wechselwirkung mit Defekten an Leerstellen geringer ist als mit Defekten, die an Zwischengitterplätze gekoppelt sind. Bei der Bildung eines molekularen Cl2 durch Reaktion zweier H- Zentren wird ein NaCI-Molekül aus seiner Gitterposition verdrängt und auf Zwischengitter-Stufenringe verdrängt, wodurch perfekte Stufenversetzungsringe entstehen.
Rekombination
Freie F- und H-Zentren rekombinieren innerhalb eines bestimmten Radius miteinander. Außerdem wird davon ausgegangen, daß
F-Zentren mit molekularen Cl2-Zentren reagieren und
H-Zentren mit F-Zentren rekombinieren können, die thermisch aus Kolloiden abgespalten worden sind.
Die erste Rekombination führt eine starke Dosisleistungsabhängigkeit auf das Kolloidwachstum ein; die beiden anderen Rekombinationsreaktionen führen dazu, daß keine Kolloidbildung bei hohen Temperaturen auftritt. Wegen der geringen Konzentrationen an S-Zentren kann die Rekombination mit F-Zentren vernachlässigt werden. Die Rekombination von H-Zentren mit F2-Zentren wird explizit berücksichtigt, die Rekombination von H-Zentren mit F3-Zentren und F4-Zentren wird über deren Bildungsgeschwindigkeiten berücksichtigt.
Die Unterschiede des Soppe-Modells zur JL-Theorie liegen
in der Akkumulation von F- Zentren in F2-, F3- und F4-Zentren und
im Einfang von H-Zentren durch Verunreinigungen, die zu S1-, S2- und S3-Zentren führen.
Der Reaktionsmechanismus, der in Bild 3 zu sehen ist, führt zu unterschiedlichen Geschwindigkeitsgleichungen für jede Defektart (siehe /SOP 93/). Einige Modellparameter wurden durch Anpassung an experimentelle Daten ermittelt. Die meisten relevanten Parameter weisen dieselben oder ähnliche Werte wie bei der JL-Theorie auf (siehe Vergleich in /GIE 93/).
Literatur
/GIE 93/ Gies, H., Hild, W., Kühle, Th. und Mönig, J.
Strahleneffekte im Steinsalz - Statusbericht
GSF-Bericht 9/93, GSF Neuherberg, (1993)
/JAI77/ Jain, U. und Lidiard, A.B.
The Growth of Colloid Centers in Irradiated Alkali Halides
Phi i. Mag. 35 (1977), 245 -259
/LID 79/ Lidiard, A.B.
Energy Stored in Irradiated NaCI
Phil. Mag. A39 (1979),647 - 659
/SEI92/ Seinen, J., Groote, J.C., Weerkamp, J.R.W. und den Hartog, H.W.
Radiation Damage in NaCI: General Model of Nucleation and Aggregation
Processes in Doped NaCI
Rad. Eff. Def. Solids 124 (1992), 325 - 339
/SOP 93/ Soppe, W.J.
Computer simulation of radiation damage in NaCI using a kinetic rate
reaction model
J. Phys.: Condens. Matter 5 (1993), 3519 - 3540
/SOP 94b/ Soppe, W.J. und Prij, J.
Kinetic model calculations of colloid growth in NaCI
Nucl. Instr. and Meth. in Phys. Res. B 91 (1994), 92 - 96
Version: 14.4.2011
Adresse dieser Seite
Home
Joachim Gruber